The endurance running hypothesis is the idea that humans evolved primarily as endurance runners. The argument goes that the human physique evolved and took its shape and function from the primary adaptive pressure of persistence hunting—that of chasing down our prey until its body shuts down.
However, this hypothesis is not without its detractors. A significant amount of scientists provide an array of counterevidence to the endurance running hypothesis. (And the debate continues.)
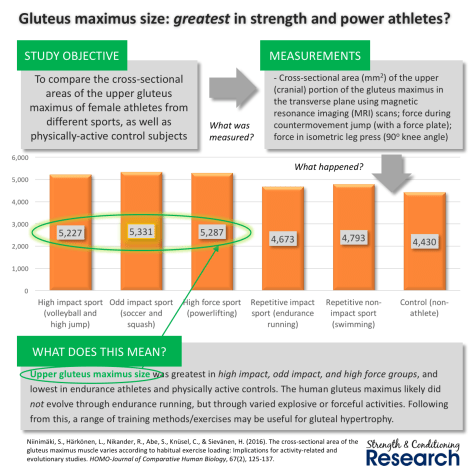
Take for example the case of the human gluteus maximus (butt muscle). Lieberman et. al. (2006) claim that the human gluteus maximus evolved its shape and size due to endurance running.
However, another article in the Journal of Comparative Human Biology finds that the gluteus maximus grows much more in high-force sports (weightlifting) and high-impact sports (such as soccer), than it does in endurance running. In fact, they also show that the butt muscle in endurance runners is no larger than in the non-athlete population.
What I disagree with is their conclusion, which is paraphrased in the “What does this mean?” section in the image below:
“The human gluteus maximus likely did NOT evolve through endurance running, but through varied explosive and forceful activities.”
My disagreements with the article (and the image) are primarily about how and why we interpret the science to mean a certain thing.
At first blush, the fact that endurance running doesn’t enlarge the gluteus maximus as much as other sports seems to detract from the idea that the muscle takes its shape from endurance running. But I think it actually adds to it.
By my analysis, these findings show that the basic, untrained shape and size of the gluteus maximus—it’s “factory specifications,” if you will—assume that it’s going to do the amounts of cutting, jumping, weightlifting, and sprinting that a habitual endurance runner might need to do. But it requires aftermarket modification to meet the (literally) outsize power and stability requirements of soccer or weightlifting.
Let’s say that a muscle evolved under a particular adaptive pressure. This means that its shape and size literally evolved to do that thing. If you take a muscle that usually doesn’t do a thing for which it evolved to do, and you ask it to do that thing, you are asking it to do something that it has prepared to do for millions of years of evolution.
In order to fit a function that it has been designed to do, the changes in shape and size that the muscle should have to undergo should be smaller, not larger. You would expect a muscle to change far more if you ask it to do something that is less aligned with its evolutionary job description.
Let’s illustrate this by looking at the arm and hand.
We probably all agree that one of the things that specifically sets us apart from our hominid cousins is the ability to coordinate the thumb with the rest of the fingers in order to grasp and manipulate objects to a high degree of dexterity. In its simplest form, this is the capability to oppose the thumb and the fingers—to make an “OK” sign with the thumb and each of the fingers of each hand.
Now let’s take a snapshot of the people who take this unique human ability to its very pinnacle: string musicians, graphic artists, etc. Their livelihood depends on the degree to which they can explore the potential of one of the major evolutionary functions of the human hand.
Compare the forearm muscles of a violinist or painter with that of a weightlifter. The weightlifter’s arms, hands, and shoulders will be much larger and more powerful. (I trust I need not cite a scientific, randomly-controlled study on the matter.) Why? Quite simple: weightlifters engage in activities that develop the body to phenomenal proportions.
But if we go by the conclusions of the article, the fact that the arm and hand get bigger through weightlifting would mean that it didn’t evolve for the kind of fine motor control that you produce in the arts. (Or that lifting heavy objects is its primary evolutionary role). A particularly ambitious version of this argument would be to suggest that one of the core functions of opposition is to become better able to lift heavy objects. But all these suppositions break down when you realize that our primate cousins were not only quite able to grasp branches and use them ably, but that opposition emerges at the same time that hominid arms were becoming smaller (and less powerful), not larger (and more powerful).
Of course, the human hand (and upper extremity in general) still needs to be able to grow and develop in order to be able to lift heavy objects—and can indeed grow to a huge degree to exhibit that function. But its core evolutionary function is to produce the unparalleled dexterity of the human being.
Furthermore, the fact that the non-painter’s hand remains relatively unchanged in size compared to the painter’s hand means that the non-painter’s hand is already relatively set up to perform that kind of dextrous function—because that’s what it presumably evolved to do. This should serve as evidence (not counterevidence) that the hand is primarily for painting (and other fine motor tasks), not for weightlifting.
We should think the same of the gluteus maximus.
Let me conclude by saying that nothing I’ve written here means that the gluteus maximus evolved exclusively for endurance running. Indeed, there is ample evidence suggesting that the architecture of the gluteus maximus is uniquely multifunction as far as muscles go. (In future posts, I’ll delve more into the nuanced view of the gluteus maximus that I proposed above: that it owes its shape and size to the fact that it is a muscle designed for the kinds of “varied explosive and forceful activities” that a bipedal, primarily endurance running animal expects to have to do.)
But what we can say is that the fact that the gluteus maximus gets bigger through a particular stimulus has no bearing on its core evolutionary role, (or on the evolutionary story of the organism as a whole).

“It is often stated that human locomotion was an adaptation to running on the open plains, which is illustrated by expressions such as ‘Savannahstan’, ‘endurance running’, ‘born to run’, ‘le singe coureur’, ‘dogged pursuit of swifter animals’ etc., even on the cover of the most influential scientific journals. Verhaegen et al. (2007) disproved in detail all endurance running arguments (Bramble & Lieberman 2004) that our Homo ancestors during most of the Pleistocene were adapted to running over open plains. When we analyse human locomotion into more elementary components, the running ‘explanation’ appears to be a just-so interpretation (cherry-picking): Bramble & Lieberman (2004) interpret every locomotor trait in humans as having evolved ‘for’ running, without even considering possible wading or swimming scenarios. A comparative approach shows that, for each trait, semi-aquatic scenarios provide more parsimonious explanations (Table 4 in Verhaegen et al. 2007), and that extant human running is a secondary and conspicuously imperfect adaptation which evolved late in the human past, for instance, we run maximally 32 km/hr over short and 20 km/hr over long distances, about half as fast as typical open plain mammals. … The nowadays popular ideas about Pleistocene human ancestors running in open plains are among the worst scientific hypotheses ever proposed. The surprising frequency and diversity of foot problems (e.g. hammertoes, hallux valgus and bunions, ingrown nails, heelspurs, athlete’s feet, corns and calluses – some of these due to wearing shoes) and the protection of our feet with shoes prove that human feet are not made in the first place for running. Moreover, humans are physiologically ill-adapted to dry open milieus: “We have a water- and sodium-wasting cooling system of abundant sweat glands, unfit for a dry environment. Our maximal urine concentration is too low for a savanna-dwelling mammal. We need more water than other primates, and have to drink more often than savanna inhabitants, yet we cannot drink large quantities at a time” (Verhaegen 1987 Nature 325:305-6). This does not imply to say that human ancestors or relatives never lived on savannas, only that if they did, it was at the wetlands and rivers there. Apparently we evolved running – only lately, and only about half as fast as equids, bovids, felids or canids, and even slower than arboreal primates – *in spite of* our broad build, short toes and plantigrade feet, profuse sweating, and large subcutaneous fat tissues (a burden of >10 kg in most people). Of course, healthy adult men can sometimes outrun ungulates (the usual ‘argument’ of conventional paleo-anthropologists) and provide a limited part of the calories for the group, but this dogged pursuit is largely confined to a few inland populations in East Africa today, is derived and probably recent (less than a few thousands of years), and it requires a rather specialized technology with water bags, weapons and poisons. Quadrupedal chimps can hunt colobus monkeys and even eat them raw, but archaic Homo with their heavy bones (POS), very broad pelves and valgus knees, shorter legs and flat feet were much too slow on land. Humans have a remarkably poor olfaction (Gilad et al. 2003) and low muscularity, which make regular scavenging, and a fortiori hunting, unlikely.” (Verhaegen 2013 Hum.Evol.28:237-266)
LikeLike
🙂 Hi Ivan, if you don’t have much time, please google
“aquatic ape theory made easy 2017”
LikeLike
Hello Marc,
I’m quite familiar with the aquatic ape theory. I hadn’t considered it previously mostly as I hadn’t ever seen it through the lens of our conversation here in the comments.
LikeLike